Natalia Pastrana-Camacho1,
Zulma Suárez1,
Alejandro Acosta-González1,
Carolina Arango1,
Brad Haltli2,
Hebelin Correa2,
Russel Kerr2,
Carmenza Duque1,
Luis Eduardo DIaz1 ![]()
For correspondence:- Luis DIaz Email: luisdb@unisabana.edu.co Tel:+5718615555
Received: 1 February 2016 Accepted: 26 May 2016 Published: 28 June 2016
Citation: Pastrana-Camacho N, Suárez Z, Acosta-González A, Arango C, Haltli B, Correa H, et al. Bioprospecting for culturable actinobacteria with antimicrobial properties isolated from rivers in Colombian Orinoquia. Trop J Pharm Res 2016; 15(6):1259-1265 doi: 10.4314/tjpr.v15i6.19
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To Isolate and characterize Actinobacteria with antimicrobial activity from Guaviare River (Colombia).
Methods: Water and sediment samples were collected from Guaviare River. Direct plating, heat and CaCO3 methods were used to isolate Actinobacteria. Six bacterial strains were tested using T-Streak method: Escherichia coli ATCC 23724, Staphylococus aureus ATCC 25923, Acinetobacter baumannii ATCC 19606, Bacillus subtilis ATCC 21556, Klebsiella pneumoniae ATCC 700603, Chromobacterium violaceum ATCC 31532. Strains of Fusarium sp. H24, Trichoderma harzianum H5 and Colletotrichum gloeosporioides were tested using Kirby-Bauer method. Isolates with high antimicrobial activity were selected for further taxonomic identification.
Results: A total of 374 actinobacteria isolates were obtained. Seven isolates exhibited high antimicrobial activity (p < 0.05) and were confirmed as members of Streptomycetaceae family. Of these, three isolates showed differential phenotypic and genotypic profiles, indicating that they may represent new species.
Conclusions: To date, this is the first study of this type in Colombian Orinoquia and indicates that this promising source of Actinobacteria from aquatic sediments with the ability to produce antimicrobial secondary metabolites.
Introduction
Actinobacteria are a predominant source of antibiotics [1,2] with Streptomyces as the major source of bioactive compounds [3]. Actinobacteria are found in soils although their isolation has focused on other unexplored habitats [2]. Aquatic habitats like rivers have been poorly explored as a source of Actinobacteria with biological activity, despite the fact that river environments can be considered a reservoir of potentially active compounds [4]. The aim of this study was to isolate and to identify Actinobacteria from water and sediments samples of Guaviare river (Colombia) and evaluated their ability to produce natural products with antimicrobial activity.
Methods
Sample collection
Water and sediment samples were collected in May 2014 from five different locations in Guaviare River (Colombia): location 1 (2°36'19.2"N 72°42'26.2"W), location 2 (2°35'24.3"N 72°41'17.2"W), location 3 (2°34'51.6"N 72°41'34.7"W), location 4 (2°33'59.4"N 72°41'43.8"W), location 5 (2°34'51.4"N 72°39'53.6"W).
Isolation of Actinobacteria
A direct plating method, a dilute heat method, and a CaCO3 enrichment method were used for isolation. For direct plating, 1 g or 1 mL of sediment/water were spread directly. For the dilute heat method 1 g or 1 mL of sediment/water was diluted in 10 mL sterile filtered river water and heated at 55 ⁰C for 6 min. For CaCO3 method 1 g of CaCO3 was mixed with 5 g sample and incubated at 45 ⁰C for 1 h [5]. Each sample from the three pretreatments was then serially diluted to 10-3 and aliquots (100 μL) were spread on oatmeal agar (ISP3), starch casein agar (SCA) and starch casein nitrate agar (SC), supplemented with cycloheximide (50 μl/mL) and nalidixic acid (10 μl/mL). Plates were incubated at 30 ⁰C for 8 days. Colonies were selected based on morphological features and were archived in 40 % (v/v) glycerol at -70 ⁰C.
Screening for antimicrobial activity
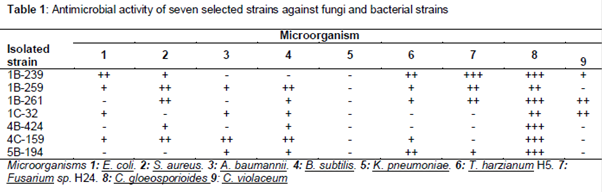
Production of antimicrobial substances and quorum sensing inhibition assay by all isolates was assessed using the T-streak method. Antibiosis was indicated by visually observable growth inhibition of the target organisms, and results were recorded as follows: ++ (complete inhibition), + (partial inhibition), – (no inhibition).
Antifungal activity
Antifungal activities were determined using the Kirby-Bauer method. Inhibition zones were recorded as follows: diameter > 20 mm: +++ (good activity), diameter between 10-20: ++ (moderate activity), diameter < 10 mm: + (weak activity) and – (no activity).
Macroscopic and microscopic features
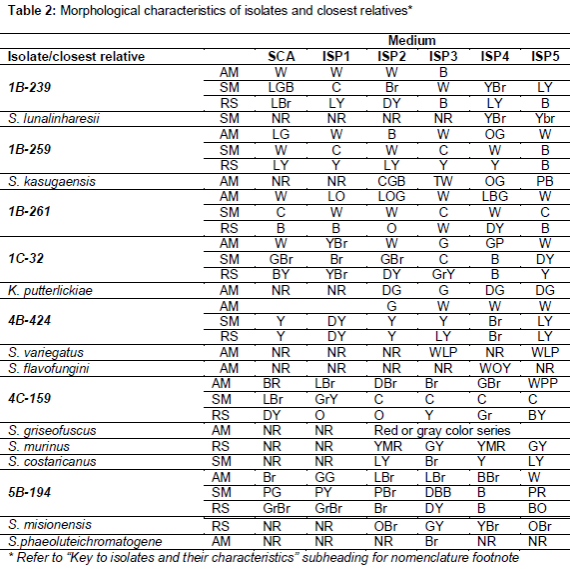
For isolates with high antimicrobial activity, different culture media were used. Media: tryptone/yeast extract agar (ISP1), malt/yeast extract agar (ISP2), oatmeal agar (ISP3), inorganic salts starch agar (ISP4), glycerol asparagine agar (ISP5) and starch casein agar SCA ().
Carbon source profiles
Isolates were grown in 12 well microplates with 5 mL basal agar [(NH4)2SO4 2.64 g/L, KH2PO4 2.38 g/L, K2HPO4 4.31 g/L, MgSO4 x 7H2O 1.0 g/L and 1.0 ml/L trace element solution (CuSO4 x 5H2O 0.64 g/L, FeSO4 x 7H2O 0.11 g/L, ZnSO4 x 7H2O 0.15 g/L, MnCl2 x 4H2O)], supplemented with 1 % (w/v) of a selected carbon source. Lactose, galactose, maltose, ribose, sucrose, xylose, inositol, fructose and carboxymethylcellulose were tested, with glucose as positive control, and a “no carbon source well”, as negative control. Growth was monitored after 5 days and results were recorded as follows: Growth equivalent to the negative control (-), growth better than negative control but not as robust as the positive control (+), growth equivalent to the positive control growth (++), growth better than the positive control (+++) ().
Taxonomical identification
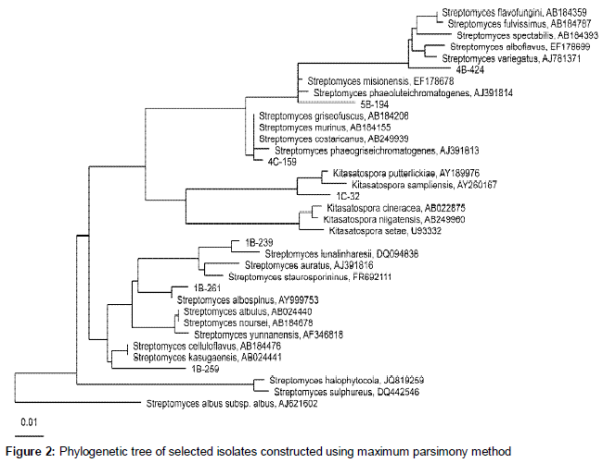
Taxonomic identification was obtained by PCR amplification, sequencing and analysis of the nearly complete 16S rRNA locus using primers 27F (5'-AGAGTTTGATCMTGGCTCAG-3') and 1492R (5'- TACGGYTACCTTGTTACGACTT-3'). PCR thermal cycling is described elsewhere [6]. PCR products were sequenced by Macrogen (Korea). Sequences were assembled using the CLC Main Workbench 7.0 program and compared with SILVA Living Tree Project (release109) database [7]. Sequences were aligned using SINA [8] and the phylogenetic tree was constructed using maximum parsimony DNA distance algorithm with ARB 5.5 [9].
Statistical analysis
The data obtained were analyzed with SAS 9.4 software using a factorial model 9x7 (nine microorganism and seven isolates) with Fisher's protected least significant difference (LSD) test for means comparison. A value of p < 0.05 was considered statistically significant.
Results
Isolation of Actinobacteria
A large number (374) of suspected Actinobacteria colonies were isolated. Pre-treatment with CaCO3, produced 275 isolates and application of the direct plating method yielded 99 isolates, whereas heat treatment did not produce isolates.
Antimicrobial activity
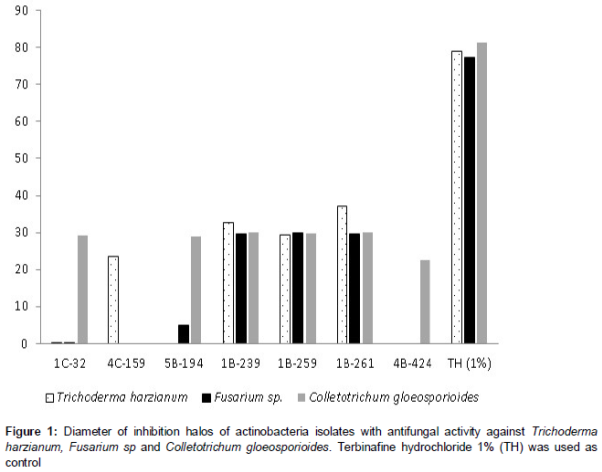
A number of 65 isolates (17.4 %) showed antibacterial activity against at least one of the bacteria tested. A significant number of isolates (149; 39.8 %) also exhibited antifungal activity to at least one of the fungus tested, showing inhibitions halos between 10 – 35 mm, corresponding to a good or moderate activity, as shown in . Statistical analysis showed an interaction between the two factors (Actinobacteria isolates and fungus strain), implying that each Actinobacteria isolate was most effective with a certain fungus strain.
Seven isolates displaying high antibacterial and antifungal activities were selected for further study. This group consisted of strains with at least moderate activity (++) or high activity (+++) against 1 fungal strain and 2 tested bacteria ().
Taxonomical identification
16S rDNA sequences were compared with SILVA Living Tree Project (release 109) database, which confirmed that all isolates belonged to Streptomycetaceae family with similarity values beyond 98 % to the closest neighbors. Maximum parsimony phylogenetic analyses is shown in .
Discussion
Enhanced recovery of Actinobacteria using the CaCO3 pretreatment may be explained by the resulting increase in pH which promotes the growth of Actinobacteria propagules [10]. Several Actinobacteria isolation protocols, utilize heating and drying stages to select heat-resistant microorganisms capable of forming spores [11]. Williams and coworkers have suggested thermal treatments between 40-50 °C for 2-16 h for selective isolation of Actinobacteria, reducing significantly accompanying flora [12]. Probably this method was not successful due the temperature used compared with exposure time.
The results suggest that all 7 strains characterized in this study belong to the Streptomycetaceae family. Isolate 5B-194 had close similarity with two Streptomyces species, but according to similarities with macroscopic characteristics (table 2) and carbon source utilization it’s probable that this isolate belongs to Streptomyces misionensis species. Isolate 4C-159 showed close similarity with three Streptomyces species, however doing the comparison between macroscopic characteristics and carbon source utilization was not possible to suggest which species belongs because differences were significant. Considering that most recent definition of a prokaryotic species based on 16S rDNA sequence similarity is determined by a minimum similarity of 98.7 % [24] strains 1B-259, 1C-32 and 4B-424 showed similarity levels to their closest neighbours below this value, suggesting that these isolates may represent new species from the Streptomycetaceae family. These strains show some differences in terms of macroscopic characteristics and carbon source utilization respect to closest neighbours. Strains 1B-239 and 1B-261, identified as Streptomyces lunalinharesii and Streptomyces albospinus, respectively. Streptomyces lunalinharesii has reported activity against Aspergillus niger, Candida albicans, Bacillus pumilus and Desulfovibrio alaskensis [13,14], as well as several phytopathogenic fungi [15]. Streptomyces albospinus produces the fungicide phenamide against Phaeosphaeria nodorum [16] and two new compounds with antibacterial activity against methicilin resistant S. aureus (MRSA) [17].
Of all isolates obtained (374 actinobacteria isolates), a large number of them were active against Bacillus subtilis, consistent with previous studies, [18] and against A. baumannii and K. pneumonia which are two carbapemen-resistant microorganisms that have become problematic due to the absence of effective medication [19]. Four strains isolated in this study showed inhibition of A. baumannii, with two of them being able to inhibit K. pneumoniae. Considering the fact that K. pneumonia used in this study is resistant to multiple antibiotics including the carbapenem-iminipem it is likely that isolates able to inhibit its growth are producers of new compounds. Our results also suggest a remarkable potential for antifungal agents, in particular against C. gloeosporioides. Fungi belonging to the genus Colletotrichum has been classified as disease-causing fungus affecting almost all agricultural crops in the world [20] as well as Fusarium spp [21]. For this reason, several actinobacterial isolates identified in this study may be potential candidates for biocontrol purposes as 43 showed inhibition of the three fungal strains tested.
Chromobacterium violaceum was included in this study to screen the strains for production of inhibitors of AHL-quorum sensing. According to these results, 257 isolates were able to inhibit pigment production in C. violaceum without altering the growth of bacteria, which represents an enormous potential, representing an alternative to the use of antibiotics and their induced resistance [22]. In this area, Actinobacteria are of interest due to their capacity to produce and secrete secondary metabolites that eventually can act as inhibitors of N-acyl-L-homoserine lactones (AHL), which mediate most Quorum sensing processes in Gram negative bacteria [23].
Conclusion
The results indicate a high abundance of culturable Actinobacteria in sediments from Guaviare River in Colombia, and also revealed a remarkable potential for 7 strains, selected as potential sources of antimicrobial compounds. Furthermore, three isolates showed differential phenotypic and genotypic profiles in comparison to those reported for the most closely related Streptomyces and Kitasatospora species, indicating they may represent new species. To the best of our knowledge, this is the first study of its type in the Colombian Orinoquía region and may help to identify new bacterial species with antimicrobial properties in this region.
Declarations
Acknowledgement
References
Archives


News Updates